अध्याय 07 मूल प्रक्रियाएँ
7.1 आनुवंशिक पदार्थ के रूप में डीएनए
आपने पिछले अध्याय में पढ़ा है कि लक्षण या गुण जीन के माध्यम से माता-पिता से संतान में वंशागत होते हैं। आप यह भी जानते हैं कि ये जीन गुणसूत्रों पर उपस्थित होते हैं जो न्यूक्लिक अम्ल और प्रोटीन से बने होते हैं। हालाँकि, उस गुण की अभिव्यक्ति के लिए उत्तरदायी जीन की प्रकृति को समझना वैज्ञानिक समुदाय के समक्ष सबसे बड़ी चुनौतियों में से एक था। इस प्रश्न का उत्तर कुछ प्रायोगिक प्रमाणों के बाद मिला कि डीऑक्सीराइबोन्यूक्लिक अम्ल (डीएनए) कुछ वायरसों को छोड़कर किसी भी जीव के लक्षण या विशेषता का निर्धारण करता है।
डीएनए की खोज का श्रेय जोहान फ्रेडरिक मीशर को जाता है, जिन्होंने पहली बार पस कोशिकाओं के केंद्रक से एक अम्लीय पदार्थ को अलग किया और न्यूक्लिन नाम दिया जिसमें डीएनए और प्रोटीन थे। गुणसूत्र और केंद्रक में उपस्थिति के कारण ये दो रासायनिक घटक; न्यूक्लिक अम्ल (मुख्यतः डीएनए) और प्रोटीन आनुवंशिक पदार्थ होने के संभावित उम्मीदवार बन गए। फिर भी, आनुवंशिक पदार्थ की प्रकृति लंबे समय तक अज्ञात रही। धीरे-धीरे, विभिन्न शोधकर्ताओं द्वारा सूक्ष्मजीवों के साथ किए गए प्रयोगों ने ऐसे परिणाम दिए जिन्होंने डीएनए को आनुवंशिक पदार्थ के पक्ष में प्रमाण प्रदान किए।
7.1.1 रूपांतरण सिद्धांत की खोज
1928 में, एक ब्रिटिश चिकित्सा अधिकारी, फ्रेडरिक ग्रिफ़िथ ने स्तनधारियों में न्यूमोनिया पैदा करने वाले जीवाणु स्ट्रेप्टोकोकस न्यूमोनिया (जिसे डिप्लोकोकस न्यूमोनिया भी कहा जाता है) के खिलाफ टीका विकसित करने के दौरान एक अवलोकन किया, जो मनुष्यों में न्यूमोनिया का कारण बनता है और आमतौर पर चूहों में घातक होता है। उन्होंने जीवाणु की दो अलग-अलग प्रजातियों (किस्मों) की पहचान की, अर्थात् विषाणुजनित (रोग उत्पन्न करने वाली) जिसमें कोशिका के चारों ओर एक पॉलीसेकेराइड कैप्सूल होता है और अविषाणुजनित (हानिरहित)। विषाणुजनित प्रजाति में, प्रत्येक जीवाणु एक पॉलीसेकेराइड कैप्सूल से घिरा होता है, जिसके कारण जीवाणु कॉलोनी जब अगर प्लेट पर उगाई जाती है तो चिकनी दिखाई देती है और इसे चिकनी प्रजाति (S) कहा जाता है। अविषाणुजनित प्रजाति में पॉलीसेकेराइड आवरण का अभाव होता है और खुरदरी दिखने वाली कॉलोनी उत्पन्न करती है और इसे खुरदरी प्रजाति (R) कहा जाता है। $\mathrm{S}$ प्रकार के जीवाणु न्यूमोनिया पैदा करके चूहों को मार देते हैं।
ग्रिफ़िथ ने $\mathrm{S}$ और $\mathrm{R}$ प्रकार के जीवाणुओं के साथ प्रयोगों की एक श्रृंखला की (चित्र 7.1)। जब उन्होंने चूहों में जीवित $\mathrm{S}$ जीवाणु इंजेक्ट किए, तो चूहों में न्यूमोनिया विकसित हो गया और वे मर गए। हालाँकि, जब उन्होंने चूहों को $\mathrm{R}$ प्रकार के जीवाणुओं से संक्रमित किया तो चूहों पर कोई बुरा प्रभाव नहीं दिखा। इन दोनों प्रयोगों के परिणामों ने इस बात की पुष्टि की कि $\mathrm{S}$ प्रकार के जीवाणुओं में मौजूद पॉलीसेकेराइड आवरण स्पष्ट रूप से विषाणुजनिता के लिए आवश्यक था।
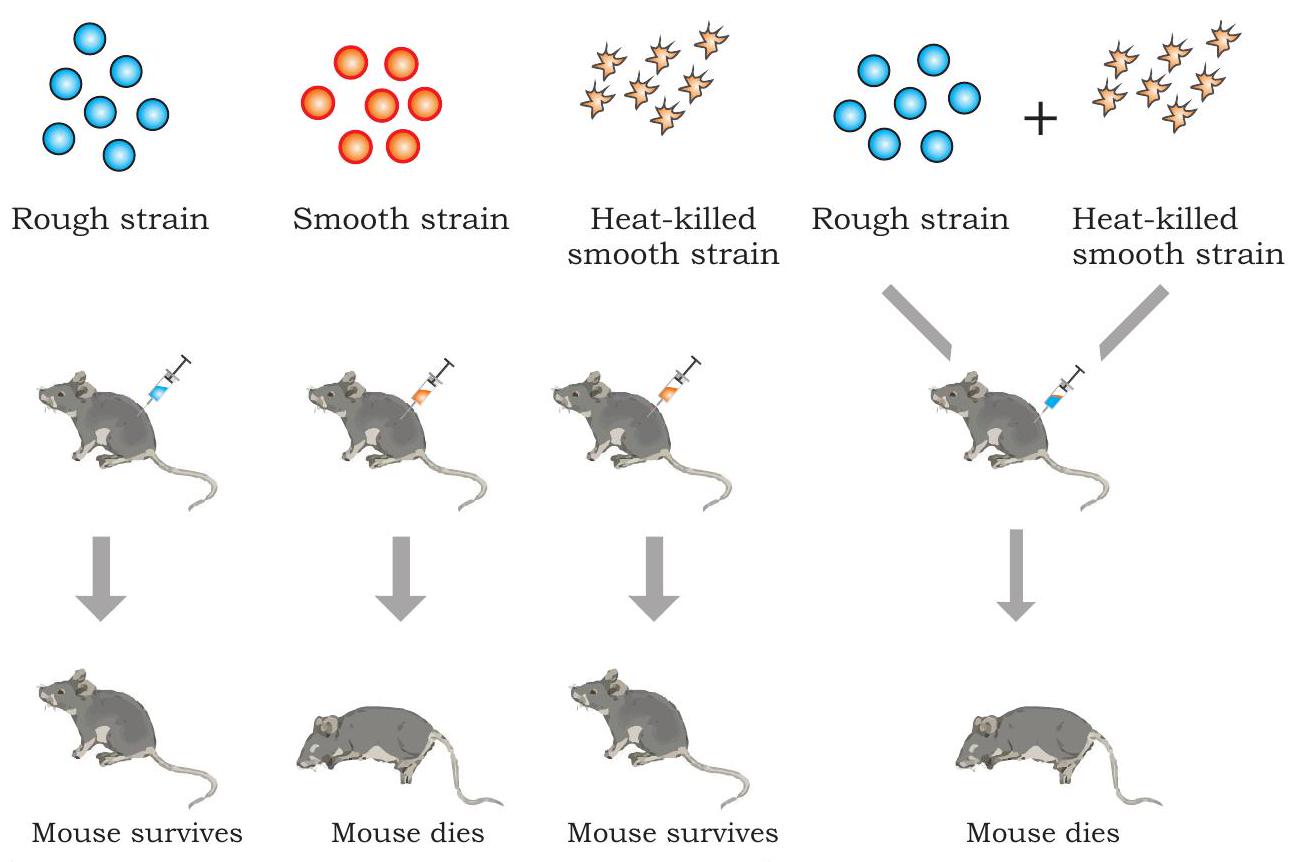
चित्र 7.1: ग्रिफ़िथ का रूपांतरण प्रयोग
आगे समझने के लिए, ग्रिफ़िथ ने कुछ विषाणुजनित $\mathrm{S}$ जीवाणुओं को उबालकर मार डाला और उक्त ऊष्मा-नष्ट जीवाणुओं को चूहों में इंजेक्ट किया। उनकी अपेक्षाओं के अनुसार, चूहे बच गए। हालाँकि, काफी अप्रत्याशित रूप से, चूहे न्यूमोनिया के कारण मर गए जब उन्हें ऊष्मा-नष्ट $\mathrm{S}$ जीवाणुओं और जीवित $\mathrm{R}$ जीवाणुओं के मिश्रण के साथ इंजेक्ट किया गया। मृत चूहों के रक्त और ऊतक द्रव की जाँच से जीवित $\mathrm{S}$ प्रकार के जीवाणुओं की उपस्थिति का पता चला। उपरोक्त अवलोकन के आधार पर, ग्रिफ़िथ ने निष्कर्ष निकाला कि R-प्रजाति के जीवाणुओं ने ऊष्मा-नष्ट $S$ जीवाणुओं से उस सिद्धांत को ग्रहण किया होगा जिसे उन्होंने ‘रूपांतरण सिद्धांत’ कहा, जिसने उन्हें चिकने-आवरण वाले जीवाणुओं में ‘रूपांतरित’ होने और विषाणुजनित बनने की अनुमति दी। उन्होंने इस घटना को रूपांतरण कहा, जिसका अर्थ है एक कोशिका से दूसरी कोशिका में आनुवंशिक पदार्थ का स्थानांतरण जो प्राप्तकर्ता कोशिका की आनुवंशिक संरचना को बदल देता है। लेकिन रूपांतरित पदार्थ की प्रकृति अभी भी निर्धारित करने की आवश्यकता थी।
7.1.2 रूपांतरण सिद्धांत की जैवरासायनिक अभिलक्षण
तीन वैज्ञानिकों, ओस्वाल्ड टी. एवरी, कोलिन मैकलियोड और मैक्लिन मैकार्टी ने ग्रिफ़िथ के रूपांतरण सिद्धांत की पहचान करने के लिए प्रयोगों की एक श्रृंखला आयोजित की, और 1944 में इसकी पुष्टि की गई कि रूपांतरण कारक डीएनए है (चित्र 7.2)। प्रयोग के डिजाइन में, उन्होंने जीवाणुओं की चिकनी प्रजाति के तीन मुख्य घटकों, यानी डीएनए, आरएनए और प्रोटीन पर ध्यान केंद्रित किया। उन्होंने जीवाणुओं की ऊष्मा-नष्ट चिकनी प्रजाति का एक अर्क तैयार किया जिसमें से लिपिड और कार्बोहाइड्रेट हटा दिए गए थे। प्रोटीन, आरएनए और डीएनए वाले अर्क के शेष घटकों को अर्क को तीन भागों में विभाजित करके आगे के प्रयोग के लिए रखा गया। इन अर्कों को अलग-अलग राइबोन्यूक्लिएज (आरएनएस), डीऑक्सीराइबोन्यूक्लिएज (डीएनएस) और प्रोटीज जैसे जलअपघटनी एंजाइमों के साथ उपचारित किया गया ताकि आरएनए, डीएनए और प्रोटीन को क्रमशः नष्ट किया जा सके, उनके रूपांतरण क्षमता के लिए एंजाइम उपचारित अर्क में से प्रत्येक को जीवाणुओं की खुरदरी प्रजाति की तीन अलग-अलग संवर्धनों में स्थानांतरित करके। खुरदरी प्रजाति का चिकनी प्रजाति में रूपांतरण उन कॉलोनियों में देखा गया जिनमें आरएनएस और प्रोटीज उपचारित अर्क मिलाया गया था और उस कॉलोनी में नहीं जिसमें डीएनएस उपचारित अर्क मिलाया गया था। इन परिणामों ने इसमें कोई संदेह नहीं छोड़ा कि यह डीएनए है जो एक संभावित रूपांतरण सिद्धांत के रूप में कार्य करता है।
परिकल्पना: कोशिकाओं का आनुवंशिक पदार्थ या तो प्रोटीन या न्यूक्लिक अम्ल (डीएनए या आरएनए) है
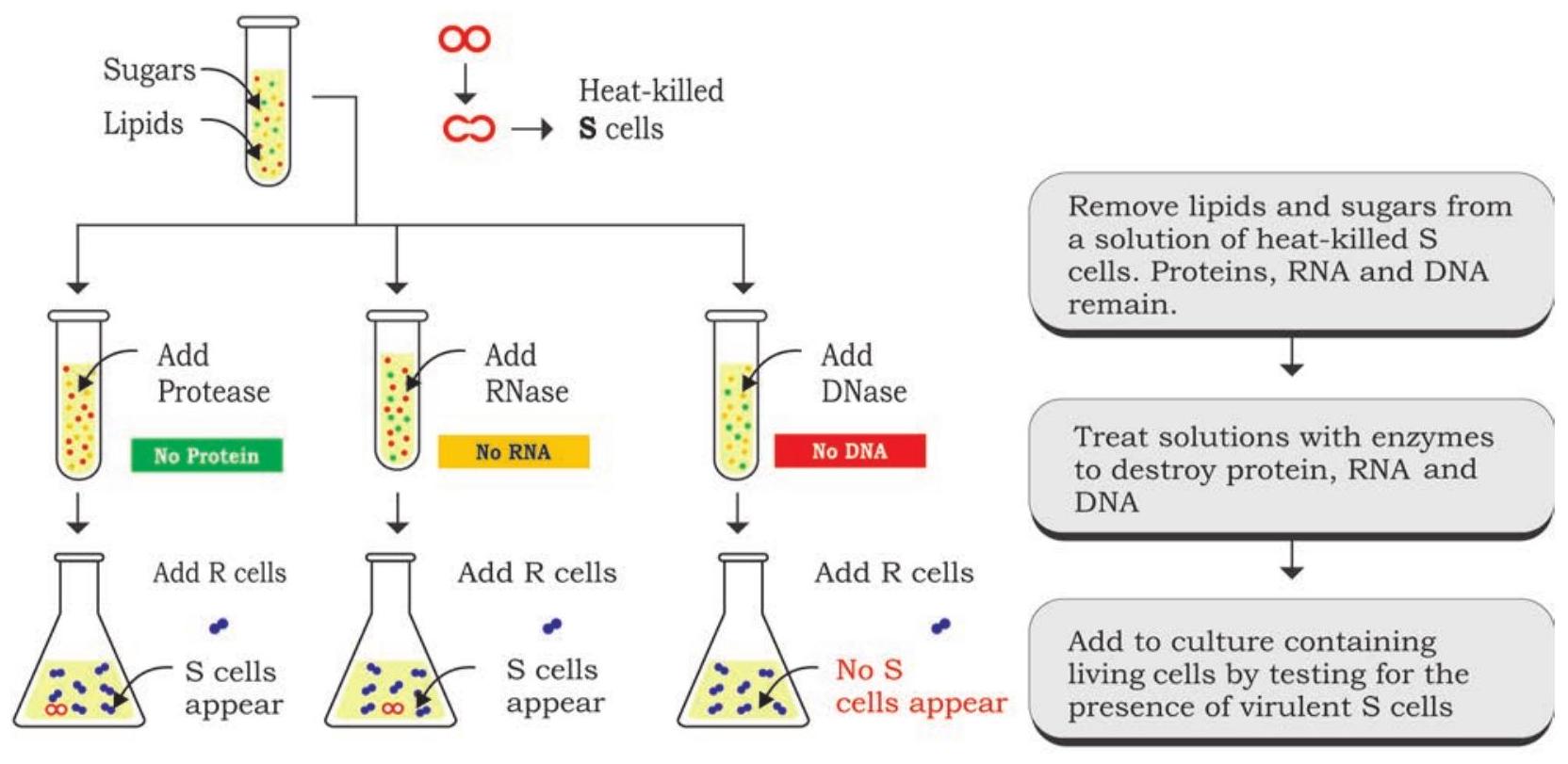
निष्कर्ष: रूपांतरण के लिए डीएनए की आवश्यकता होती है, इसलिए यह कोशिका का आनुवंशिक पदार्थ है
चित्र 7.2: रूपांतरण सिद्धांत की पुष्टि
7.1.3 हर्शे - चेस प्रयोग
बाद में, अल्फ्रेड हर्शे और मार्था चेस (1952) द्वारा T2 बैक्टीरियोफेज के साथ किए गए एक और प्रयोग ने डीएनए को आनुवंशिक पदार्थ के पक्ष में प्रमाण प्रदान किया। T2 बैक्टीरियोफेज वायरस जो एस्चेरिचिया कोलाई जीवाणु को संक्रमित करता है, उसमें प्रोटीन आवरण से घिरा डीएनए होता है। जब यह एक जीवाणु कोशिका को संक्रमित करता है, तो यह बाहरी सतह से जुड़ जाता है और फिर अपने डीएनए को कोशिका में इंजेक्ट कर देता है। T2 बैक्टीरियोफेज और E. कोलाई के साथ उनके प्रयोगों की एक श्रृंखला में, उद्देश्य यह स्थापित करना था कि कौन सा घटक फेज कणों के गुणन के लिए उत्तरदायी है, डीएनए या प्रोटीन। आसानी से पहचान करने के लिए, $\mathrm{T} 2$ बैक्टीरियोफेज को शुरू में $E$ कोलाई की कॉलोनियों के साथ अलग-अलग रेडियोधर्मी फॉस्फोरस $\left({ }^{32} \mathrm{P}\right)$ और रेडियोधर्मी सल्फर $\left({ }^{35} \mathrm{S}\right)$ युक्त माध्यम में उगाया गया (चित्र 7.3)। इसके परिणामस्वरूप बैक्टीरियोफेज के एक समूह को रेडियोधर्मी फॉस्फोरस $\left({ }^{32} \mathrm{P}\right)$ से और दूसरे समूह को रेडियोधर्मी सल्फर $\left({ }^{35} \mathrm{S}\right)$ से चिह्नित किया गया।
${ }^{35} \mathrm{S}$ और ${ }^{32} \mathrm{P}$ चिह्नित $\mathrm{T} 2$ फेज को अब अचिह्नित $E$ कोलाई जीवाणु कॉलोनी की दो अलग-अलग संवर्धनों में इनोकुलेट किया गया। संक्रमण के बाद, जीवाणु कॉलोनियों को जीवाणु कोशिकाओं के बाहर से किसी भी शेष फेज और फेज भागों को हटाने के लिए एक ब्लेंडर में हिलाया गया। ब्लेंडर के मिश्रण को फिर जीवाणुओं (पेलेट में मौजूद) को फेज अवशेषों (सुपरनेटेंट में मौजूद) से अलग करने के लिए अपकेंद्रित किया गया। जीवाणु संवर्धन के पेलेट जिन्होंने रेडियोधर्मिता दिखाई, वे रेडियोधर्मी डीएनए वाले फेज से संक्रमित थे, जबकि, सुपरनेटेंट में रेडियोधर्मिता देखी गई जो ${ }^{35} \mathrm{S}$ बैक्टीरियोफेज से संक्रमित थी। यह इंगित करता है कि प्रोटीन फेज से जीवाणुओं में प्रवेश नहीं करते थे। इसलिए, यह निष्कर्ष निकाला गया कि वह पदार्थ जो जीवाणु कोशिका में प्रवेश करता है, यानी डीएनए आनुवंशिक पदार्थ हो सकता है।
हालाँकि उपरोक्त प्रयोगों ने डीएनए को आनुवंशिक पदार्थ के पक्ष में मजबूत प्रमाण प्रदान किए, लेकिन यह स्पष्ट नहीं था कि डीएनए अणु आनुवंशिक सूचना का भंडार है। एरविन चारगाफ, मॉरिस विल्किंस, रोज़लिंड फ्रैंकलिन, जेम्स वाटसन और फ्रांसिस क्रिक द्वारा किए गए बाद के अध्ययनों के कारण डीएनए संरचना की खोज हुई, जिससे यह स्पष्ट हुआ कि डीएनए बड़ी मात्रा में सूचना को कैसे संकेतित कर सकता है (इकाई II, अध्याय 3 में वर्णित)।
$\hspace{3.5cm}$बैक्टीरियोफेज

चित्र 7.3: हर्शे-चेस प्रयोग
7.2 प्रोकैरियोटिक और यूकैरियोटिक जीन संगठन
यह अच्छी तरह से समझा जाता है कि गुण माता-पिता से संतान में ‘जीन इकाई’ के रूप में वंशागत होते हैं और डीएनए कुछ वायरसों को छोड़कर सभी जीवों में आनुवंशिक पदार्थ है (जहाँ आनुवंशिक पदार्थ आरएनए है)। इससे जीन के संगठन के बारे में प्रश्न उठा, क्या यह संगठन प्रोकैरियोट्स के साथ-साथ यूकैरियोट्स में भी समान है और यह आणविक स्तर पर कैसे कार्य करता है? जीन वंशागति की इकाई है जो एक विशिष्ट गुण या लक्षण को नियंत्रित करती है और वैकल्पिक रूपों में भी व्यक्त हो सकती है जिन्हें एलील कहा जाता है। दूसरे शब्दों में जीन डीएनए का एक खंड है जो आरएनए संश्लेषण के माध्यम से पॉलीपेप्टाइड श्रृंखला के संश्लेषण के माध्यम से स्वयं को व्यक्त करता है, जिसे आणविक जीव विज्ञान का ‘केंद्रीय सिद्धांत’ माना जाता है।
शुरुआत में यह स्थापित किया गया था कि गुण या लक्षण 1900 में मेंडल के वंशागति के सिद्धांत की पुनर्खोज के आधार पर जीन द्वारा नियंत्रित होते हैं, और पौधों और जानवरों दोनों में बाद के शोधों की एक श्रृंखला ने इस तथ्य की स्थापना की कि लक्षण या विशेषताएँ कुछ अंतर्निहित सिद्धांतों द्वारा नियंत्रित और विनियमित होते हैं जो एक पीढ़ी से दूसरी पीढ़ी में पारित होते हैं। गुण या लक्षण को नियंत्रित करने वाले कारक या अंतर्निहित इकाई को बाद में 1909 में विल्हेम जोहानसेन द्वारा ‘जीन’ नाम दिया गया।
जीन की प्रकृति और कार्यप्रणाली को समझना बीसवीं शताब्दी की शुरुआत के दौरान वैज्ञानिक समुदाय के मुख्य फोकस में से एक था। 1930 के दशक के दौरान जॉर्ज बीडल और एडवर्ड टैटम द्वारा कवक न्यूरोस्पोरा क्रासा पर किए गए कार्य ने एक एंजाइम के संश्लेषण को नियंत्रित करने के लिए एक जीन के संबंध को स्थापित करने में मदद की।
इस गुण पर विचार करते हुए कि कवक न्यूरोस्पोरा क्रासा को सरल शर्करा, अकार्बनिक लवण और विटामिन बायोटिन युक्त माध्यम पर बहुत आसानी से उगाया जा सकता है, बीडल और टैटम ने इस आधार पर प्रयोग किया कि जीव अन्य आवश्यक अमीनो अम्ल और नाइट्रोजनी क्षारकों का स्वयं संश्लेषण कर सकता है (चित्र 7.4)। इसे स्पष्ट माना गया कि संश्लेषण एंजाइमों द्वारा मध्यस्थ होता है, जो आनुवंशिक नियंत्रण के तहत संश्लेषित होते हैं। प्रयोग का डिजाइन बहुत सरल था जिसमें कोनिडिया, यानी अलैंगिक बीजाणुओं को उत्परिवर्तन प्रेरित करने के लिए एक्स-रे के साथ विकिरणित किया गया था (चित्र 7.4)। विकिरणित बीजाणुओं द्वारा उत्पादित संतानों की पहचान कुछ विशिष्ट न्यूनतम माध्यम पर उगाकर की गई। उत्परिवर्तन वाली प्रजातियों की पहचान करने के उद्देश्य से, विकिरणित बीजाणुओं की संतानों को जंगली प्रकार के साथ संकरित किया गया और बाद की संतानों को किसी विशिष्ट अमीनो अम्ल या विटामिन के लिए न्यूनतम माध्यम पर उगाकर उत्परिवर्तित प्रजातियों की पहचान की गई (ऐसा संवर्धन माध्यम जिसमें एक विशिष्ट विटामिन या अमीनो अम्ल को छोड़कर सभी अमीनो अम्ल, नाइट्रोजनी क्षारक और विटामिन होते हैं)। उनके द्वारा ऐसे कई उत्परिवर्तनों की पहचान की गई और यह आनुवंशिक रूप से स्थापित किया गया कि प्रत्येक उत्परिवर्तन, वास्तव में, एक विशिष्ट एंजाइम के गैर-कार्य करने के परिणामस्वरूप होता है।
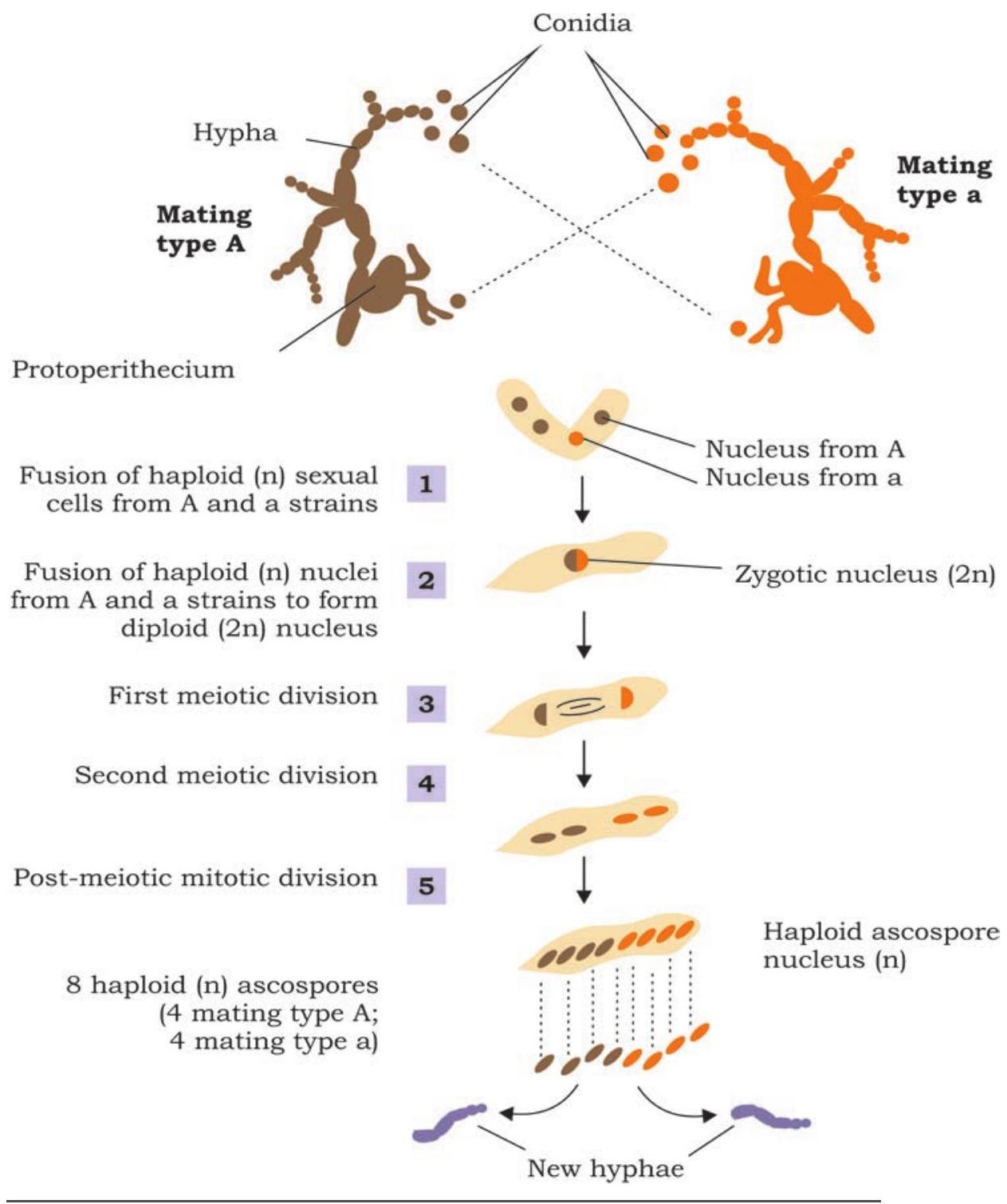
चित्र 7.4: न्यूरोस्पोरा में उत्परिवर्तन का पता लगाने वाला प्रयोग
बाद में, यह देखा गया कि सभी प्रोटीन एकल पॉलीपेप्टाइड से नहीं बने होते हैं बल्कि एक से अधिक पॉलीपेप्टाइड श्रृंखला से बने होते हैं। इस तथ्य के कारण कि एक जीन एक पॉलीपेप्टाइड को संकेतित करता है; केंद्रीय सिद्धांत भी एक जीन एक प्रोटीन से संशोधित होकर एक जीन एक पॉलीपेप्टाइड हो गया।
लगभग एक साथ 1940 के दशक की शुरुआत में, क्रोमैटिन तंतु की कोशिका विज्ञानी जाँच ने इलेक्ट्रॉन सूक्ष्मदर्शी के माध्यम से कुछ हद तक धागे पर मनके जैसी संरचना प्रकट की (चित्र 7.5) और यह आसानी से निष्कर्ष निकाला गया कि प्रत्येक मनका शायद एक जीन का प्रतिनिधित्व करता है।

चित्र 7.5: क्रोमैटिन की धागे पर मनके संरचना
बाद की जाँचों से पता चला कि प्रत्येक मनका एक न्यूक्लियोसोम है (जिसमें हिस्टोन ऑक्टामेर का एक कोर और $146 \mathrm{~bp}$ का द्वि-रज्जुक डीएनए होता है) और दो मनकों के बीच का धागा, लिंकर डीएनए। यह भी स्थापित किया गया कि प्रत्येक न्यूक्लियोसोम अपने लिंकर क्षेत्र के साथ लगभग $200 \mathrm{~bp}$ शामिल करता है। इसे एक जीन नहीं माना जा सकता, क्योंकि कई मामलों में जीनों का आकार 200 न्यूक्लियोटाइड से बहुत बड़ा होना चाहिए। सरल कारण यह है कि कई प्रोटीनों में 100 से अधिक अमीनो अम्ल अवशेष होते हैं और उनके संबंधित नियामक जीन उसी के तीन गुना से कम नहीं हो सकते (कोडॉन की त्रिक प्रकृति के आधार पर)।
अब यह स्पष्ट हो गया है कि एक जीन विशिष्ट प्रोमोटर क्षेत्र वाला डीएनए का खंड है, जहाँ आरएनए पॉलीमरेज़ बंध सकता है और एमआरएनए का अनुलेखन कर सकता है। अनुलेखित एमआरएनए फिर अनुवाद की प्रक्रिया में शामिल हो जाता है। यह तंत्र वायरस से लेकर जीवाणु, पौधों और जानवरों तक सभी जीवों के लिए समान है। क्या वायरस, जीवाणु या उच्च जीव के जीन उनकी संरचना और कार्य में समान हैं? गुणसूत्रों के एक पूर्ण समुच्चय (सभी जीनों और डीएनए के अन्य भागों सहित) में कुल डीएनए सामग्री को जीनोम कहा जाता है। जैसा कि हम वायरस या जीवाणु के मामले में देखते हैं, जीनोम का आकार तुलनात्मक रूप से यूकैरियोटिक जीनोम से बहुत छोटा होता है। यूकैरियोटिक जीनोम प्रोकैरियोटिक जीनोम की तुलना में बहुत अधिक जटिल होते हैं। पादप जीनोम किसी भी अन्य यूकैरियोटिक जीनोम की तुलना में और भी अधिक जटिल होते हैं। जीवित जीवों के विभिन्न समूहों में जीनोम के अनुमानित आकार को चित्र 7.6 में दिखाया गया है।
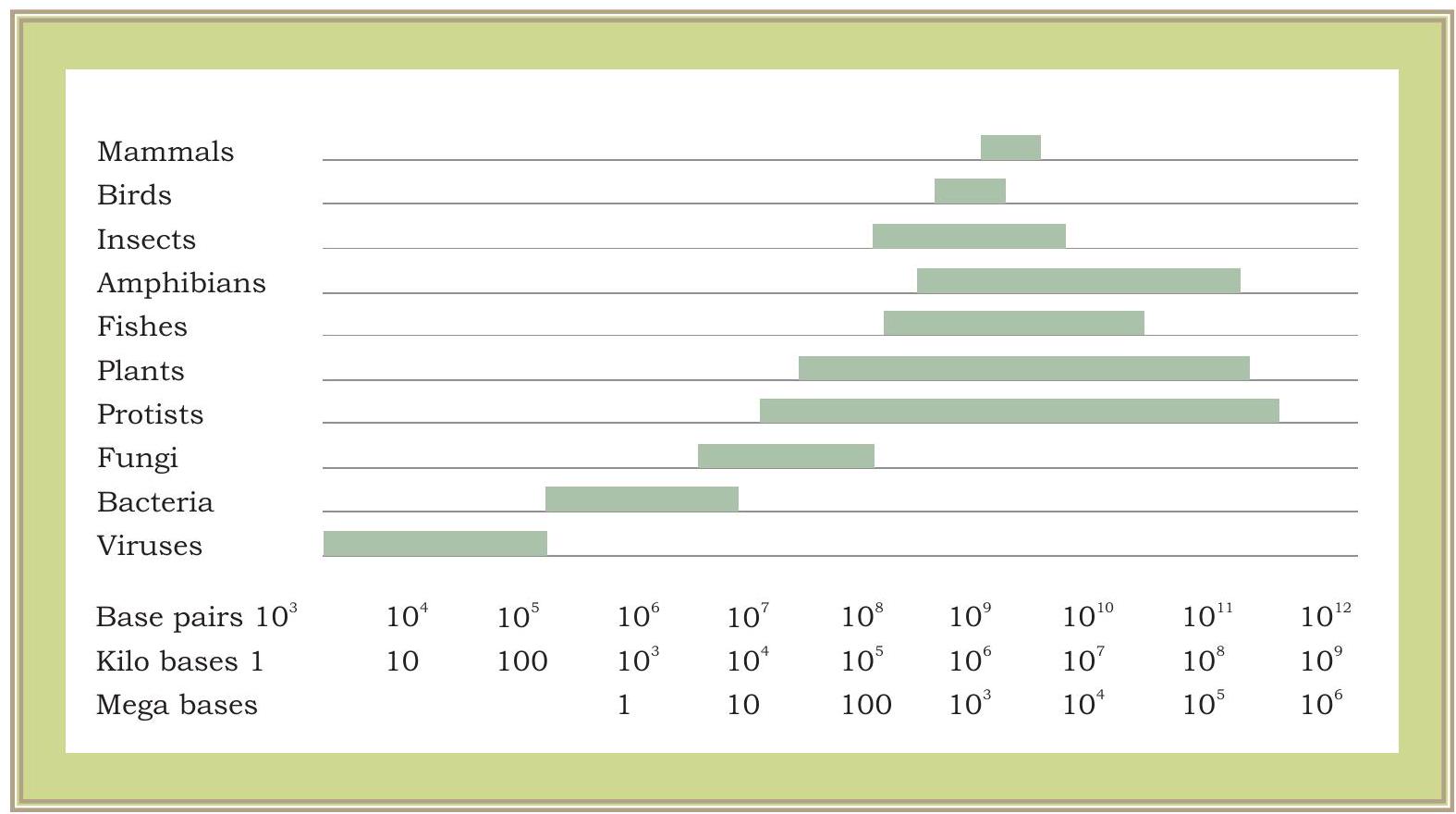
चित्र 7.6: जीवित जीवों के विभिन्न समूहों में जीनोम आकार में भिन्नता। जीनोम आकार हजारों न्यूक्लियोटाइड युग्मों में मापा जाता है, यानी $1000 \mathrm{bp}=1$ किलोबेस (kb) और 1000,000 $\mathrm{bp}=1000 \mathrm{~Kb}=1$ मेगाबेस (Mb)
अधिकांश यूकैरियोट्स में जीनोम का एक बड़ा हिस्सा व्यक्त नहीं होता है और गैर-कोडिंग अनुक्रम के रूप में रहता है। यह भी देखा गया है कि यूकैरियोटिक जीन अभिव्यक्ति में पॉलीपेप्टाइड श्रृंखला संश्लेषण की प्रक्रिया में शामिल सक्रिय एमआरएनए का आकार प्राथमिक प्रतिलेख से बहुत छोटा होता है। वास्तव में, कई यूकैरियोटिक जीन, उदाहरण के लिए, हीमोग्लोबिन का $\beta$-ग्लोबिन जीन अनुलेखन के बाद स्प्लाइसिंग की प्रक्रिया से गुजरता है जिसमें प्राथमिक प्रतिलेख के कुछ बिखरे हुए खंडों को हटा दिया जाता है (इंट्रॉन) और आरएनए प्रतिलेख के शेष भागों (एक्सॉन) को एक साथ जोड़कर एमआरएनए बनाया जाता है।
एक यूकैरियोटिक कोशिका में दो प्रकार के जीनोम होते हैं: (i) केंद्रकीय जीनोम और (ii) अंगकीय जीनोम।
केंद्रकीय जीनोम
डीएनए का अधिकांश भाग केंद्रक में पाया जाता है और इसे केंद्रकीय डीएनए के रूप में जाना जाता है। प्रोकैरियोट्स में, अधिकांश जीनोम कोडिंग डीएनए अनुक्रमों से बना होता है जबकि यूकैरियोटिक जीनोम में कोडिंग क्षेत्र कुल जीनोम का अपेक्षाकृत बहुत छोटा हिस्सा बनाते हैं। उदाहरण के लिए, मानव जीनोम का आकार लगभग $3,000 \mathrm{Mb}$ या 3 अरब क्षारक युग्म डीएनए का है और अनुमानित रूप से 20,000 से अधिक जीन हैं, जो कुल जीनोम का लगभग $2 %$ बनाते हैं। जीनोम के गैर-कोडिंग क्षेत्र में, ऐसे अनुक्रम होते हैं जो टैंडेम सरणियों के रूप में हजारों से कई मिलियन बार दोहराए जाते हैं। जीनोम में इन दोहराए जाने वाले डीएनए अनुक्रमों का आकार और संख्या काफी भिन्न होती है।
एक यूकैरियोटिक कोशिका में केंद्रकीय जीनोम को छोटे संघनित इकाई में संगठित किया जाता है, जिसे रेखीय डीएनए अणु वाले गुणसूत्र के रूप में जाना जाता है। एक व्यक्ति के अगुणित जीनोम में गुणसूत्रों का एक पूर्ण समुच्चय अक्षर n द्वारा दर्शाया जाता है। अधिकांश जीव प्रत्येक कोशिका में गुणसूत्रों के दो समुच्चय रखते हैं और इन्हें द्विगुणित जीव (2n) के रूप में जाना जाता है।
अंगकीय जीनोम
केंद्रकीय डीएनए के अलावा, कुछ झिल्ली-बद्ध कोशिकीय अंगक जैसे हरितलवक और सूत्रकणिका में अंगक-विशिष्ट डीएनए होता है। अंगक जीनोम ज्यादातर वृत्ताकार द्वि-रज्जुक डीएनए होते हैं और प्रत्येक अंगक में एकाधिक प्रतिय